Throughout life the human brain is constantly changing and modifying itself to adapt to the changing demands of the environment. This so-called cognitive plasticity is an inherent characteristic of the brain that essentially reflects one’s ability to learn. Older individuals tend to vary substantially to the degree that they show improvement through cognitive intervention and therefore document varying degrees of cognitive plasticity. Indeed, older individuals tend to be characterised by great variability in terms of how much they may benefit from mental training.
This underscores the need to understand both the brain mechanisms that predispose older individuals to benefit greatly from mental training, as well as the brain mechanisms underlying the inability to benefit from such training.
In our latest research paper we conceptualised cognitive plasticity as the extent to which older individuals benefited from a memory training intervention.
For some years our group has been running an extensive training study (see NCP project) where our participants have undergone either intensive memory training to remember word lists, or been assigned to a control group that received no specific memory training. During memory training participants learnt to use a technique known as the Method of Loci, whereby in order to memorize a list of objects in sequential order one must visualise each of the objects and mentally place them along a familiar route. By this method, each object to-be-remembered becomes associated with a particular location along a journey, and this enables easier recall.
Participants underwent MRI scans at the onset of the intervention, and every ten weeks during the study.
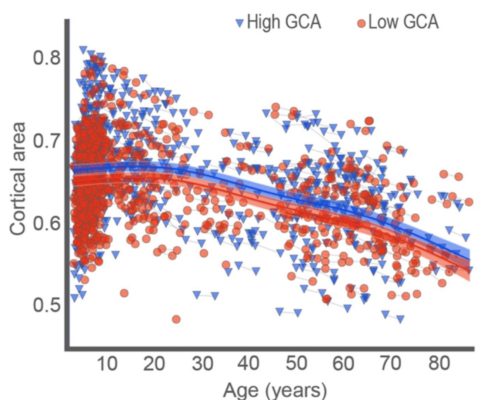
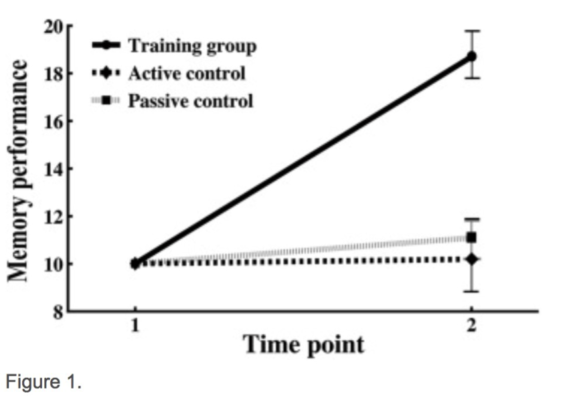
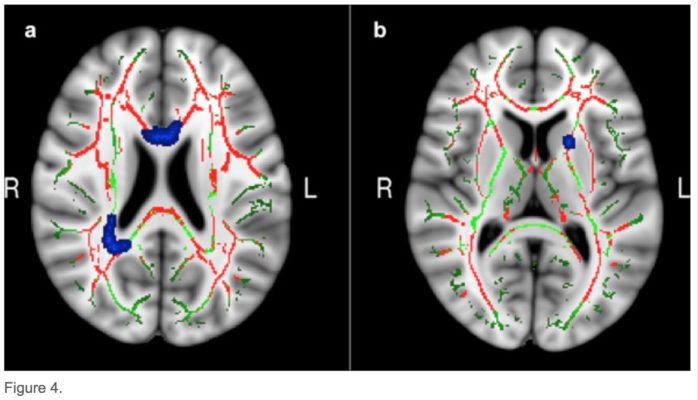
As predicted, only the group that underwent memory training had shown an improvement in memory score by the end of the study period (above figure), demonstrating our memory-training paradigm to be effective. In the brain’s white matter tracts it was found that relative to older adults, younger adults tended to show a greater degree of white matter integrity (as inferred by diffusion measurements of molecular motion) across many regions of the brain. In addition, the analysis led to the identification of 3 specific areas of white-matter that related to the degree of memory improvement from the training we administered (below figure).
This was the key finding, as it indicates that individuals with a lager degree of white matter microstructural integrity as measured before the training period, benefitted more from the training and thus were able to improve their memory scores to a greater extent.Therefore the integrity of the brain’s white matter tracts could be a potential marker for one’s ability to adapt in response to cognitive demands during ageing.
Citation
98. White-matter integrity as a marker for cognitive plasticity in ageing 
Neurobiology of Aging
Glasø de Lange, A; Bråthen, A. C; Grydeland, H; Sexton, C; Johansen-Berg, H; Andersson, J; … Walhovd, K. (2016)